




Introduction
Successful implantation requires a competent blastocyst, a receptive endometrium, and adequate crosstalk between the two. In humans, fecundability is poor (20-25%), and approximately 75-80% of pregnancy failures result from implantation failure [1]. The human endometrium undergoes dynamic changes, with menstrual breakdown and subsequent regeneration during each menstrual cycle [2]. Intricate changes at the tissue, cellular, and molecular levels during each menstrual cycle are required to create a period of receptivity for blastocyst implantation [3,4]. This period of receptivity is known as the “window of implantation (WOI)”, a term first used by Edwards to describe the human uterus [5]. The WOI is a short period that begins on day 19 or 20 of the menstrual cycle and persists for 4-5 days [2,6].
Since histological assessment of the endometrium is insufficient to predict the receptivity for embryo implantation [7], investigations have focused on examining the molecular physiology of the WOI in the human endometrium. Consequently, several biomarkers have been suggested as uterine receptivity markers including hormone receptors, biochemical markers, and immunohistochemical markers [8,9]. Given the complexity of endometrial-regenerating tissues and recent advances in microarray technology, researchers have begun to use transcriptomics to characterize endometrial changes [4,10], especially for assessing endometrial receptivity [11]. The endometrial receptivity array (ERA) is a bulk-tissue transcriptomic analysis method that involves the retrieval of a small amount of tissue from endometrial biopsies in the WOI. This array has been used to schedule embryo transfers in clinically selected women undergoing in vitro fertilization and embryo transfer (IVF-ET) [12,13]. The ERA is a customized microarray containing 238 genes that are differentially expressed during different phases of the menstrual cycle. This transcriptomic signature allows prediction of WOI displacements and identification of the endometrial receptivity status as “receptive” or “non-receptive”, findings that can be used to guide personalized embryo transfer (pET) [12].
Nevertheless, questions remain regarding the clinical benefits of ERA and whether ERA testing increases the success rate of pET. Some studies have shown higher rates of implantation and ongoing pregnancy in patients who underwent ERA-based management [14,15], while others have not [16-18]. Although ERA testing significantly increased the cumulative pregnancy rate in a randomized controlled trial of patients with infertility, it did not change the first-attempt success rate when compared with standard procedures involving transfers of fresh or thawed embryos [19]. Emerging evidence has shown that pET based on ERA does not improve pregnancy rates and may negatively affect reproductive outcomes [20]. These negative effects have been attributed to the presence of more variations in transcriptomic endometrial signatures than previously reported, with ERA testing leading to inaccurate recommendations regarding embryo transfer, as well as harmful progesterone exposure because of the increased duration before embryo transfer [20].
As studies on the transcriptomic signatures of the WOI have reported differing results [21], further transcriptomic research evaluating the optimal endometrial receptivity is warranted. Biological processes and molecular participants in the transition from the early secretory (ES) phase to the mid-secretory (MS) phase of the endometrium underscore the complex events involved in preparation for embryo implantation. Previously reported genes with altered expression during the endometrial transition from the ES to the MS phase were frequently involved in ion binding, cell cycle regulation, transport of signaling proteins, and immune modulation [22]. Although several studies have compared gene expression in the receptive (i.e., MS) and pre-receptive (ES) phases in an attempt to identify the molecular signature characteristic of a receptive endometrium [22-25], no consensus has been reached regarding the genes accounting for transcriptomic differences between phases.
Therefore, we conducted this study to evaluate endometrial transcriptomic patterns in the ES and MS phases of the natural menstrual cycle before IVF-ET and to compare differences in differentially expressed genes (DEGs) according to subsequent IVF-ET outcomes (pregnancy or no pregnancy).
Materials and methods
1. Study population
Thirty patients aged 29-40 years who underwent IVF-ET were included in this study. To exclude embryonic effects on implantation, only patients who underwent the transfer of ≥1 high-quality embryo were included. Embryos with ≥6 blastomeres (development grade I/V or II/V on day 3) were considered to be morphologically of high quality. Patients with adenomyosis, submucosal or endometrial-distorting uterine myomas, endometrial hyperplasia, endometrial cancer, endometrial polyps, inflammatory diseases, infectious diseases, malignancy, cardiovascular diseases, or autoimmune diseases were excluded from this study.
Endometrial samples were obtained from the included patients using a Pipelle catheter (Cooper Surgical, Trumbull, CT, USA) during the ES or MS phases of the natural menstrual cycle immediately before IVF-ET. Endometrial dating was histologically confirmed as the ES phase (days 16-18 of the menstrual cycle) or the MS phase (days 19-21 of the menstrual cycle), according to the Noyes criteria. We divided the patients into two groups depending on the IVF-ET outcome: pregnant group (n=14; 7 in ES and 7 in MS) and non-pregnant group (n=16; 8 in ES and 8 in MS).
This study was approved by the Institutional Review Board of Severance Hospital (IRB No. 4-2018-0747), and informed consent was obtained from all participants.
2. Ribonucleic acid (RNA) extraction and quantification
RNA was extracted from formalin-fixed paraffin-embedded (FFPE) tissues. The samples were cut into sections with a thickness of 10 μm, and five sections were deparaffinized with xylene and rehydrated with ethanol. RNA was extracted using a High Pure FFPET RNA Isolation Kit (Roche, Mannheim, Germany) according to the manufacturer’s instructions. RNA purity and concentration were measured using a NanoDrop (Thermo Scientific, Waltham, MA, USA), and RNA integrity was assessed using an RNA 6000 Nano Chip on an Agilent 2100 Bioanalyzer (Agilent, Santa Clara, CA, USA).
3. Gene expression quantification and differentially expressed gene analysis
RNA extracted from FFPE samples was analyzed using the nCounter® analysis system (NanoString Technologies, Inc., Seattle, WA, USA) in accordance with the manufacturer’s instructions. A total of 5 μL of each RNA sample was combined with 8 μL of Codeset Master Mix containing the Reporter CodeSet and hybridization buffer. We then added 2 μL of the capture probe set, after which the contents were mixed by vortexing for 10 seconds and briefly spun down. Next, the solution was placed in a thermal cycler (Bio-Rad Laboratories Inc., Hercules, CA, USA) for 16 hours, after which the samples were moved to the prep station containing the prepared cartridges and the nCounter Master Kit. At this station, the samples were purified and bound to a cartridge. This process was completed in approximately 2.5 hours, with 12 lanes per run. The cartridges were then transferred from the prep station to a digital analyzer (NanoString Technologies), where they were scanned using a setting of 555 fields of view.
To quantify gene expression values, first, a normalization step was performed using the geNorm algorithm [26] in the nCounter Advanced Analysis version 2.0.115 (NanoString Technologies® Inc., nSolver™ Advanced Analysis Software [NanoString Technologies, Inc.]; https://www.nanostring.com/products/analysis-software/advanced-analysis). Second, the genes that were differentially expressed in the two selected biological environments were analyzed using the default option. To compare gene expression profiles, nCounter Advanced Analysis version 2.0.115 clustered the normalized gene expression values of a few hundred selected DEGs in an unsupervised manner. The software generated expression plots for each gene, as well as volcano plots showing the magnitude of change and P-values for comparisons between the two samples.
To understand the biological functions of the DEGs, we performed functional enrichment analysis using g:Profiler [27] and cluster Profiler [28]. This involved performing gene-set overlap testing and a comparison of the analyzed DEGs and a number of functionally categorized genes, including biological processes of Gene Ontology, Kyoto Encyclopedia of Genes and Genomes (KEGG) pathways, and other functional gene sets.
4. Statistical analysis
Statistical analyses were performed using Statistical Package for the Social Science, version 26.0 (SPSS Inc., Chicago, IL, USA). Patient characteristics were compared between the pregnant and non-pregnant groups using Fisher’s exact test for categorical variables and the Mann-Whitney-Wilcoxon test for continuous variables. Statistical significance was set at P<0.05.
nSolver software (freely available from NanoString Technologies) was used for mRNA data analysis. The lists of upregulated and downregulated genes were obtained and filtered using the criterion of a 2-fold change to identify DEGs. Differences in gene expression were assessed using the Mann-Whitney-Wilcoxon test, with P-values <0.05 indicating statistical significance.
Results
1. Clinical characteristics in study population
There were no significant differences in the age, parity, duration of infertility, number of previous IVF attempts, or cause of infertility between the pregnant and non-pregnant groups (Table 1).
2. Gene expression profiles of the early secretory and mid-secretory phase endometria in the pregnant group
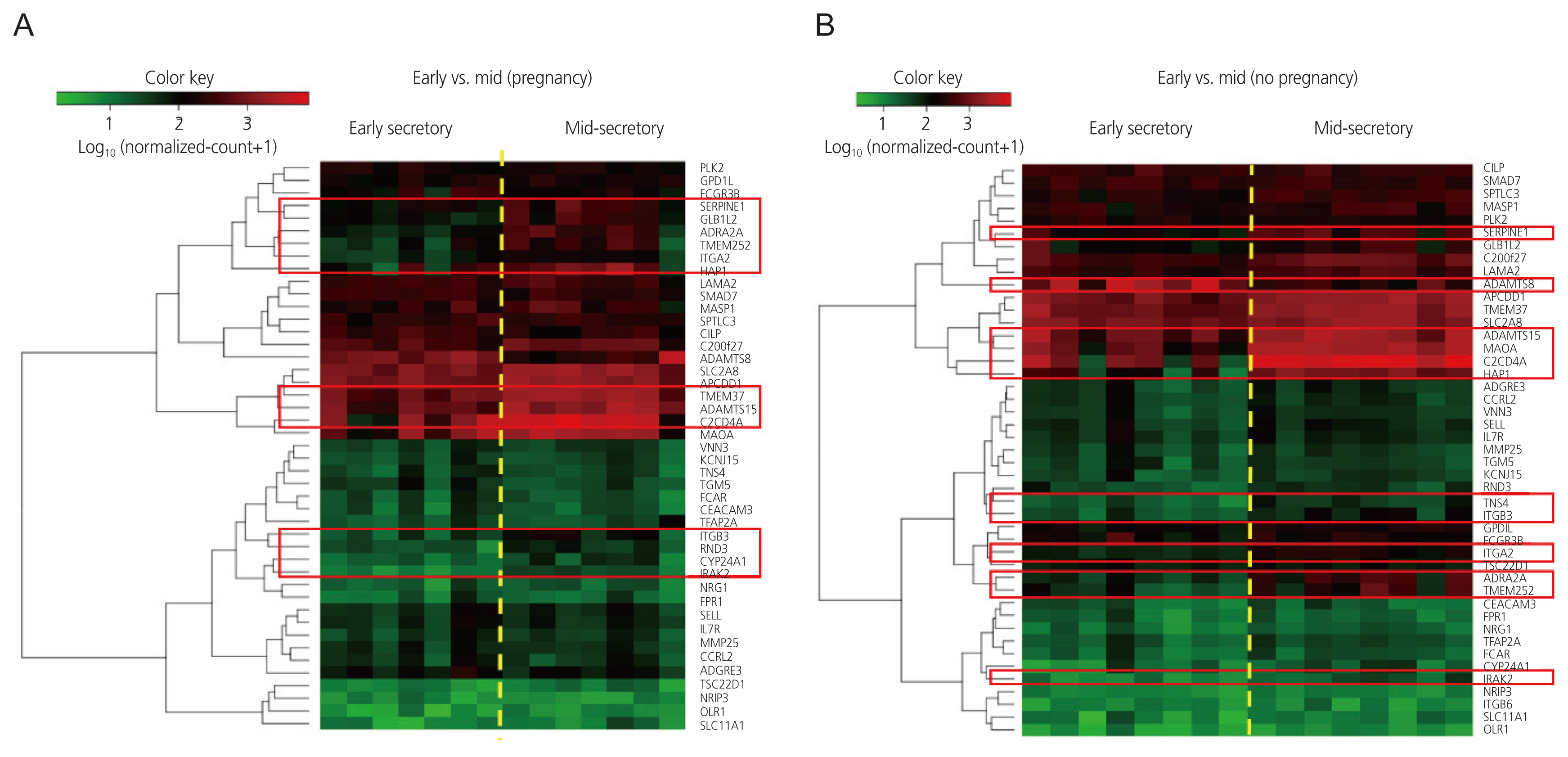
Heatmaps generated after raw data analysis revealed distinct expression profiles of upregulated genes in the MS phase endometrium compared to the ES phase endometrium in the pregnant group (Fig. 1A). The volcano plot (Fig. 2A) shows all samples plotted as a function of fold change versus P-value. Thirteen DEGs were identified when comparing the ES and MS phase endometria in the pregnant group (Table 2).
3. Gene expression profiles of the early secretory and mid-secretory phase endometria in the non-pregnant group
Heatmaps generated after raw data analysis revealed distinct expression profiles of both upregulated and downregulated genes in the MS phase endometrium versus the ES phase endometrium in the non-pregnant group (Fig. 1B). The volcano plot (Fig. 2B) shows all samples plotted as a function of fold change versus P-value. A total of 11 DEGs were identified when comparing the ES and MS phase endometria in the non-pregnant group (Table 3).
4. Comparison of gene expression patterns between pregnant and non-pregnant groups
No DEGs were found between the pregnant and non-pregnant groups in the ES and MS phases (data not shown). Nine genes were upregulated in the MS phase endometrium, compared to the ES phase endometrium, in both the pregnant and non-pregnant groups: adrenoceptor alpha 2A, interleukin 1 receptor-associated kinase 2, a disintegrin and metalloproteinase with thrombospondin repeats 15, serpin family E member 1, integrin subunit beta 3, transmembrane protein 252, huntingtin associated protein 1 (HAP1), C2 calcium-dependent domain containing 4A and integrin subunit alpha 2. In contrast, there were differences in the expression patterns of several genes between the ES and MS phases in the pregnant and non-pregnant groups. Four genes were upregulated in the MS phase endometrium, compared to the ES phase endometrium, though this was only in the pregnant group: transmembrane protein 37 (TMEM37), galactosidase beta 1 like 2 (GLB1L2), Rho family GTPase 3 (RND3), and cytochrome P450 family 24 subfamily A member 1 (CYP24A1). One gene (ADAMTS8) was downregulated and one gene (MAOA) was upregulated in the MS phase endometrium, compared to the ES phase endometrium, though this was only in the non-pregnant group. The molecular functions, biological processes, and KEGG pathways of the DEGs, with patterns that differed between the pregnant and non-pregnant groups, are shown in Table 4.
Discussion
This study showed that in a natural menstrual cycle before IVF-ET, endometrial transcriptomic patterns differed between the ES and MS phases according to IVF-ET outcomes. We identified 9 DEGs with upregulated expression in the MS phase endometrium (compared to the ES phase endometrium), regardless of whether pregnancy occurred. In contrast, TMEM37, GLB1L2, RND3, and CYP24A1 were upregulated only in the pregnant group, ADAMTS8 was downregulated, and MAOA was upregulated only in the non-pregnant group. Among these selected DEGs, some were previously reported, while others were novel.
The function of the TMEM37 protein has not yet been validated, but it may inhibit calcium influx through a subunit of calcium channels and may be related to insulin release [29]. It may also be associated with the prognosis of patients with colon cancer [30]. The GLB1L2 protein is a galactosidase with beta-galactosidase activity that is involved in carbohydrate metabolic processes [31]. However, its physiological function remains largely unknown.
The RND protein is a member of the Rho family, a group of proteins that was first characterized by their ability to induce loss of cell-substrate adhesion, followed by cell rounding through the inhibition of actin stress fiber formation in fibroblasts [32]. RND has multiple regulatory functions in carcinogenesis [33], cardiovascular disease [34], preeclampsia [35], and neural development [36]. Its upregulation would likely increase myosin phosphatase activity, which enhances uterine relaxation during pregnancy [37].
Vitamin D is mainly synthesized in the skin and, to a lesser extent, is obtained from food sources. It is a key modulator of Ca and phosphorus regulation. Vitamin D is first transported to the liver, where it is converted to 25-hydroxyvitamin D (25(OH)D3) in the liver mitochondria. It is further metabolized to the active form of vitamin D, 1,25-dihydroxyvitamin D (1,25(OH)2D3), in a process that occurs mainly in the kidneys [38]. The human endometrium expresses CYP24A1 either constitutively [39] or after stimulation with 1,25(OH)2D3 [40]. Thus, vitamin D3 signaling may participate in the normal functioning of the uterus and placenta during the pre- and post-implantation periods.
Monoamine oxidases are key regulatory molecules in monoamine neurotransmitter metabolic pathways and include the MAOA and monoamine oxidase B (MAOB) isoenzymes, which differ in their substrate selectivity and inhibitor sensitivity. Both MAOA and MAOB are located on the X chromosome but encode different proteins [41]. In recent years, investigators have suggested that MAOA may play an important role in promoting tumor migration and invasion, epithelial cell proliferation, and epithelial-mesenchymal transition [42]. Although these data suggest that the role of MAOA is not limited to the nervous system, its contribution to endometrial receptivity remains unclear. In a previous study of genes at different phases of the endometrial cycle, which included an analysis of published DNA sequencing ChIP data, MAOA was found to be highly expressed in the MS phase endometrium, compared to the ES phase endometrium, whereas the expression of MAOB did not change significantly between the two phases [4]. These microarray results suggested that MAOA may be involved in the establishment of endometrial receptivity. Previous experimental results have also revealed that deficient MAOA expression in the endometrium is associated with implantation failure in recipients of donated oocytes [43]. In women with adenomyosis and in an adenomyotic mouse model, reduced MAOA activity impaired endometrial receptivity by promoting inappropriate proliferation of endometrial epithelial cells via downregulation of the forkhead box protein O1 during the WOI [44]. In contrast, our results showed that MAOA was upregulated in the MS phase (compared with the ES phase) only in the non-pregnant group.
ADAMTS is a metalloproteinase family of secreted zinc-dependent proteases composed of 19 genetically distinct members in humans [45]. Most ADAMTS subtypes play a critical role in the remodeling of the extracellular matrix, and their expression is related to prenatal and postnatal growth as well as the development of several diseases, including cancer, arthritis, Alzheimer’s disease, and a number of inflammatory and thrombotic conditions [45-47]. Due to the ability of some ADAMTS proteins to cleave large chondroitin sulfates, subtypes 1, 4, 5, 8, 9, and 15 have been classified as aggrecanases [46]. The expression and function of aggrecanases in steroid-mediated extracellular matrix remodeling of the endometrium remain poorly characterized; however, it has been suggested that gonadocorticoids exhibit complicated regulatory activities on the expression of aggrecanases in primary cultures of human endometrial stromal cells. ADAMTS subtypes may play crucial roles in endometrial physiology [48].
HAP1, the first protein shown to interact with the huntingtin protein, is specifically expressed in human brain tissue and is involved in the pathogenesis of Huntington’s disease [49]. HAP1 is also found in the pituitary gland, thyroid gland, adrenal medulla, pancreatic islets, and other endocrine organs and tissues [50]. A previous study showed that HAP1 is involved in vesicular transport [51]. Therefore, HAP1 may regulate macromolecule secretion in the secretory endometrium. However, it was previously reported that HAP1 mRNA expression in the endometrium on menstrual cycle day 21 did not differ between patients with idiopathic infertility and fertile control patients [52].
Our study has several limitations. First, the sample size was relatively small. Therefore, our results should be considered preliminary and further large-scale studies are warranted. Secondly, because endometrial tissue samples were obtained from the menstrual cycle before IVF-ET, and not the actual IVF-ET attempt cycle, they may not reflect changes in an actual pregnancy cycle. However, it is not ethically possible to retrieve endometrial tissue during an IVF-ET attempt cycle because the sampling process may be detrimental to pregnancy outcomes. In addition, ovarian stimulation may affect endometrial receptivity compared to a natural cycle, suggesting that this may be due to supraphysiological concentrations of steroids during controlled ovarian stimulation [53]. Several studies have investigated the effects of different stimulation cycles on the endometrial receptivity transcriptomics. In previous studies that focused on the secretory phase, it was generally observed that while the expression patterns in the pre-receptive phase were similar between natural and stimulated cycles, notable differences were found when comparing WOI [54-57]. Therefore, further studies are necessary to evaluate endometrial transcriptomic patterns in natural menstrual cycles or hormone therapy cycles before frozen-thawed embryo transfer cycles, and to compare differences in DEGs according to embryo transfer outcomes. Nonetheless, the DEGs identified in the present study may be different from those in the subsequent IVF-ET cycle, and we believe that these novel DEGs may play a role in endometrial receptivity. Third, embryonic effects on implantation may have been unaccounted for. Although only patients with morphologically high-quality embryos(s) were included in our study, we did not screen for chromosomal abnormalities that may have affected pregnancy outcomes. Therefore, further investigations using euploid embryos are required. Fourth, we included endometrial samples obtained in the ES (days 16-18 of the menstrual cycle) or MS (days 19-21 of the menstrual cycle) phases. Considering that the endometrium changes rapidly around the time of implantation, the inclusion of samples collected on different menstrual cycle days could introduce significant variance in the results.
In conclusion, changes in endometrial transcriptomic patterns from the ES phase to the MS phase in a natural menstrual cycle immediately before IVF-ET appear to differ according to pregnancy outcomes. These novel DEGs, which have not been previously studied, may have functional significance during WOI and serve as potential biomarkers of endometrial receptivity.







")











