




Introduction
Cervical cancer, comprising 80-85% squamous cell carcinoma (SCC) and 15-20% adenocarcinoma (AC), is caused by high-risk human papillomavirus (HR HPV) infection [1,2]. Integration of HR HPV DNA into the genome of cervical epithelial cells causes genome instability and leads to altered cellular processes and signaling pathways such as phosphatidylinositol-3 kinase (PI3K)/Akt, Wnt/╬▓-catenin, Raf/MEK/extracellular signal-regulated kinases (ERK), apoptosis and coupled membrane receptor signaling [3,4,5,6].
Glycogen synthase kinase 3╬▓ (GSK3╬▓) is the key enzyme for multiple signaling pathways including PI3K/Akt and Wnt/╬▓-catenin and its activity is modulated by site-specific phosphorylation [7]. GSK3╬▓ is present in an activated form, GSK3╬▓ phosphorylated at Tyr216 (pY216GSK3╬▓), or in an inactivated form, GSK3╬▓ phosphorylated at Ser9 (pS9GSK3╬▓), and the expression levels of these forms vary depending on the tumor site and type [8,9,10,11,12].
Cervical SCC and AC share common etiologic and risk factors such as the total number of sexual partners of the patient and the age at first intercourse. However, with respect to smoking, long duration smoking is a stronger risk factor for SCC than for AC [2]. Although there are few studies describing common alterations in the signaling pathways in cervical AC and SCC, it remains unclear whether there are substantial differences in the molecular pathology involved in the development of each tumor type [3].
In our previous work, we demonstrated that the expression of GSK3╬▓ increases with disease progression from cervical intraepithelial neoplasia (CIN) to invasive cancer, and the expression of GSK3╬▓ was higher in SCC than in AC, suggesting the possible involvement of GSK3╬▓ in the development of SCC [13]. However, only a small number of AC cases was included and there were no data on the expression of the phosphorylated forms of GSK3╬▓ (pY216GSK3╬▓ and pS9GSK3╬▓) in that study.
In this study, we expanded on the observations of the previous study by performing immunohistochemical staining for pY216GSK3╬▓ and pS9GSK3╬▓ as well as GSK3╬▓ in additional cases of SCC and AC, and we compared their expression patterns between the 2 tumor types to identify their significance in SCC and AC development.
Materials and methods
1. Specimens
A total of 84 cervical cancer specimens (64 SCC and 20 AC) with 62 CIN (31 CIN1 and 31 CIN3) and 12 normal cervical tissues were used in this study. To perform immunohistochemical staining for GSK3╬▓, pY216GSK3╬▓, and pS9GSK3╬▓ in the CIN, SCC, and AC tissues, we used the previously constructed tissue microarrays (TMAs) containing 62 CINs, 56 SCCs, and 7 ACs [13] as well as a newly constructed TMA containing 8 SCCs and 13 ACs. In case the tissues are lost in TMAs as they have been reused from the previous study, corresponding whole sections from the representative blocks were prepared. The HPV data of the patients were not available in most of the cases; therefore, we used p16 immunostaining as an ancillary test for a marker of HPV infection [14].
2. Immunohistochemistry
Immunohistochemical staining was performed using an autostainer (DakoCytomation, Carpinteria, CA, USA). Four-micrometer-thick tissue sections were obtained from the TMA blocks and mounted on poly-L-lysine coated slides. After deparaffinization and rehydration, antigen retrieval was performed by heating the sections in citrate buffer (pH 6.0) at 121┬░C for 10 minutes. Endogenous peroxidase activity was blocked with 3% hydrogen peroxide for 5 minutes, and the sections were incubated with primary antibodies against GSK3╬▓ (BD Biosciences, Lexington, KY, USA; 1:250), pS9GSK3╬▓ (Abcam, Cambridge, UK; 1:250), pY216GSK3╬▓ (Abcam; 1:250), and p16 (p16INK4a kit). Color development and counterstaining of the sections were performed by diaminobenzidine and hematoxylin. Tissue sections from glioblastoma multiforme were used as positive controls for GSK3╬▓ and those from pancreatic and colonic AC were used as positive controls for pS9GSK3╬▓ and pY216GSK3╬▓, respectively [15,16]. The sections that were not incubated with the primary antibodies served as negative controls. GSK3╬▓, pS9GSK3╬▓, and pY216GSK3╬▓ expression was considered as positive when more than 10% of the tumor area showed nuclear and/or cytoplasmic or membranous staining with any intensity. The expression of p16 was considered as positive when p16 showed strong and diffuse block-positivity in the tumor cells [17].
3. Statistical analysis
The association between pY216GSK3╬▓, GSK3╬▓, and pS9GSK3╬▓ expression and clinicopathological characteristics, such as age, parity, tumor size, the International Federation of Gynecology and Obstetrics (FIGO) stage, lymphovascular space invasion (LVSI), lymph node (LN) metastasis, and pathologic type was analyzed using chi-square or Fisher's exact test. Comparative analysis of immunoexpression between normal ectocervix, CIN, and invasive cancer (SCC and AC) was also performed using chi-square and Fisher's exact test. Comparative analysis of immunoexpression between pY216GSK3╬▓, GSK3╬▓, and pS9GSK3╬▓ in SCC and AC was performed using McNemar test. In all the experiments, a P-value <0.05 was considered statistically significant. Statistical analysis of the data was performed using the SPSS ver. 21.0 (SPSS Inc., Chicago, IL, USA).
Results
The age of the cancer patients ranged from 27 to 81 years and the mean age was 49.6 years. Out of the 84 invasive cervical cancer patients, 37 cases were classified as stage IA, 27 as stage IB, 5 as stage II, 12 as stage III, and 3 as stage IV of cervical cancer, according to FIGO staging system. The patient characteristics and the expression patterns of GSK3╬▓, pS9GSK╬▓, and pY216GSK3╬▓ based on each characteristic are summarized in Table 1. Significant associations were observed between protein expression and the patient's age, parity, and tumor types (SCC and AC), but no correlations was found between the expression of these 3 proteins with other characteristics such as tumor size, FIGO stage, LVSI, and LN metastasis.
Table┬Ā1
Analysis of pY216GSK3╬▓, GSK3╬▓, and pS9GSK3╬▓ expression in cervical carcinoma with clinicopathological characteristics
Values are presented as number (%). The Žć2 or Fisher's exact test. Bold values denote statistical significance at the P<0.05 level.
FIGO, International Federation of Gynecology and Obstetrics; LVSI, lymphovascular space invasion; LN, lymph node; SCC, squamous cell carcinoma; AC, adenocarcinoma.
The expression levels of pY216GSK3╬▓, GSK3╬▓, and pS9GSK3╬▓ in normal cervix, SCC, and AC are shown in Fig. 1. Non-neoplastic ectocervical squamous cells showed strong staining for pY216GSK3╬▓ (12/12, 100%) and negative staining for GSK3╬▓ (0/12, 0%) and pS9GSK3╬▓ (0/12, 0%). In the endocervix, pY216GSK3╬▓ staining was weakly positive, but GSK3╬▓ and pS9GSK3╬▓ staining was negative in the columnar cells lining the endocervical surface with the exception of positive staining for pS9GSK3╬▓ in endocervical glands. In CIN, GSK3╬▓ expression (18/62, 29%) was increased in comparison to the normal ectocervix (P=0.0318) but there was no significant difference in the expression of pY216GSK3╬▓ (58/62, 94%) and pS9GSK3╬▓ (13/62, 21%) between CIN and the normal ectocervix, respectively (P>0.9999, P=0.1098).
Fig.┬Ā1
Result of pY216GSK3╬▓, GSK3╬▓, and pS9GSK3╬▓ expression in normal cervix, squamous cell carcinoma (SCC), and adenocarcinoma (AC).
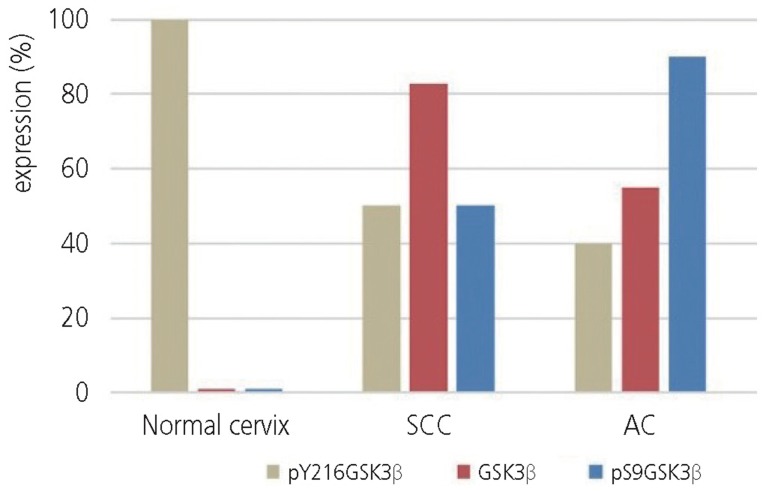
In SCC, the expression levels of GSK3╬▓ (53/64, 83%) (P<0.0001) as well as pS9GSK3╬▓ (32/64, 50%) (P=0.0013) were significantly increased compared to that in the normal ectocervix. However, the expression level of pY216GSK3╬▓ in SCC (32/64, 50%) was significantly lower compared to that in the normal ectocervix (P=0.0013). In AC, the expression levels of pS9GSK3╬▓ (18/20, 90%) and GSK3╬▓ (11/20, 55%) were significantly higher (P<0.0001, P=0.0016) while pY216GSK3╬▓ expression level (8/20, 40%) was lower (P=0.0006) compared to that of normal endocervix. The comparative analysis of GSK3╬▓, pS9GSK3╬▓, and pY216GSK3╬▓ expression revealed that GSK3╬▓ was significantly increased compared to pS9GSK3╬▓ or pY216GSK3╬▓ in SCC, whereas pS9GSK3╬▓ was significantly increased compared to GSK3╬▓ or pY216GSK3╬▓ in AC (Table 2).
Table┬Ā2
Comparative analysis of pY216GSK3╬▓, GSK3╬▓, and pS9GSK3╬▓ expression in squamous cell carcinoma and adenocarcinoma
Values are presented as number (%). McNemar test. Bold values denote statistical significance at the P<0.05 level.
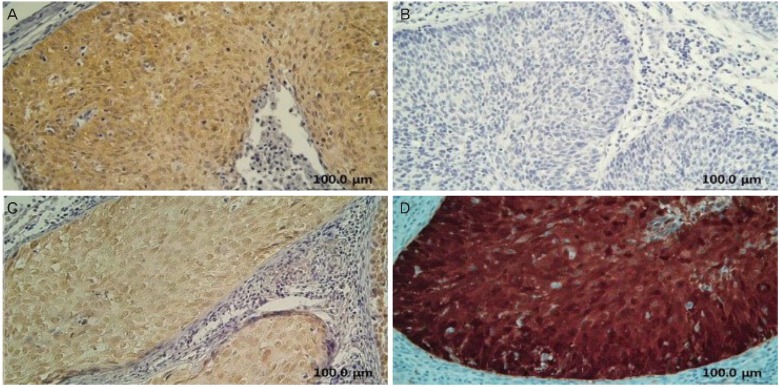
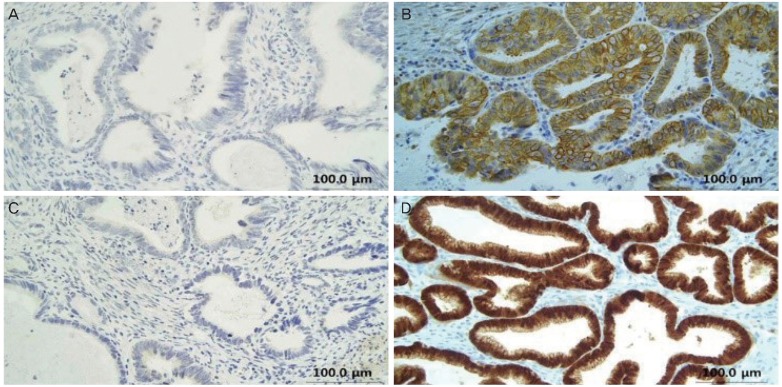
Further, the examination of the subcellular localization of GSK3╬▓, pS9GSK3╬▓, and pY216GSK3╬▓ showed that these three proteins were localized mostly in nucleus and/or cytoplasm of SCC cells (Fig. 2). However, in AC cells, pS9GSK3╬▓ showed additionally membranous localization whereas GSK3╬▓ and pY216GSK3╬▓ showed nuclear and/or cytoplasmic localization (Fig. 3). The p16 was localized in both nuclear and cytoplasmic in all of the SCC and AC cases (Fig. 2D and 3D).
Discussion
In the present study, the expression patterns of GSK3╬▓/pS9GSK3╬▓ and pY216GSK3╬▓ were inversely proportional in cervical cancer; higher expression of GSK3╬▓ and pS9GSK3╬▓ and lower expression of pY216GSK3╬▓ was found in both SCC and AC compared to the normal cervix, which is consistent with the previous results [9,13]. In cervical SCC, decreased expression of pY216GSK3╬▓ is directly associated with loss of adenomatous polyposis coli (APC) and inversely with nuclear ╬▓-catenin, suggesting that the reduction of pY216GSK3╬▓ may occur simultaneously with the disruption of GSK3╬▓-axin destruction complex by Wnt activation [9]. It has been reported that HPV E6 and E7 interact with p53 and Rb, leading to loss of cell cycle control [18] and participate in regulation of Wnt/╬▓-catenin pathway [9,19]. In addition, overexpression of pS9GSK3╬▓ and c-Myc in CIN and SCC, compared to the normal tissues, is significantly associated with HPV16 infection, suggesting the downstream effects of PI3K/Akt signaling in cervical cancer [9]. PI3K/Akt pathway is frequently activated in cervical neoplasia by gain-of-function mutation of PIK3CA resulting in Akt phosphorylation [3,20,21]. Previously, we have shown that PI3K-p110╬▒ overexpression and its positive correlation with pAkt in CIN3 and SCC which supports the possible involvement of PI3K/Akt activation in cervical carcinogenesis [21]. Similar results were obtained during mouse skin tumorigenesis; Tyr216 dephosphorylation and Ser9 phosphorylation occur in cancer cells from Akt-transformed mouse keratinocytes [22].
Despite increased pS9GSK3╬▓ expression, GSK3╬▓ was the most elevated component in cervical SCC. It is largely unknown how GSK3╬▓ level is elevated in tumor cells, but GSK3╬▓ overexpression in cervical cancer appears to be determined at the genomic level [23]. Genome wide expression analysis shows up-regulation of genes (GSK3╬▓, FZD2, PPAR╬┤ and c-Myc) related to the Wnt/╬▓-catenin signaling pathway in HPV16-positive cervical SCC, which is validated by in situ hybridization [23]. Simultaneous elevation of GSK3╬▓ and pS9GSK3╬▓ in the same tumor cells may suggest that the activity of GSK3╬▓ in Wnt pathway is independent of pS9GSK3╬▓ in PI3K/Akt pathway [24]. GSK3╬▓ overexpression has been also reported in tumors other than cervical cancers [8,10,13,25]. Aberrant nuclear accumulation of GSK3╬▓ is associated with tumor dedifferentiation in pancreatic AC and urothelial carcinoma of the bladder [25,26]. GSK3╬▓ also plays a role in cancer progression by participating in nuclear factor kappa B (NF-╬║B)-mediated gene transcription and modulating apoptotic pathway [27,28].
In this study, although pY216GSK3╬▓ expression was decreased in cervical cancer compared to the normal cervix, its level was maintained in about half of SCC and AC cases. This implicates that a basal level of pY216GSK3╬▓ activity is still required in cancer cells similar to the resting cells, where the expression of pY216GSK3╬▓ is constitutive [29].
It is well known that GSK3╬▓ is largely present in the cytoplasm and also within nuclei and mitochondria, but there is little information about the subcellular localization of pS9GSK3╬▓ [12,30,31]. In neuronal cells, pS9GSK3╬▓ is implicated in axonal growth via actin polymerization and microtubule assembly which is induced by local nerve growth factor mediated PI3K activation [32]. The pS9GSK3╬▓ is also involved in the branching of the podocyte processes and elongation by microtubule polymerization and stabilization in compensatory glomerular adaptation to podocyte loss [33]. In an earlier study, we have shown that pS9GSK3╬▓ was located on the membrane, with/without cytoplasmic localization, of normal acinar or ductal cells, co-localized with cytokeratin 7 [15]. These results suggest that pS9GSK3╬▓ plays a role in cellular outgrowth and morphologic changes in various cell types by regulating or collaborating with cytoskeletons [33].
In the present study, pS9GSK3╬▓ was predominantly localized in the membrane of AC cells, whereas it was present in the nucleus and/or cytoplasm of SCC cells, suggesting a different role of pS9GSK3╬▓ depending on cell type and subcellular localization. The membrane localization of pS9GSK3╬▓ in glandular cells may be related to Akt activation-mediated morphogenesis [34]. In mammary cells, Akt activation elicits large misshapen structures that cooperate with oncoproteins such as cyclin D1 or HPV E7 [34]. In lung carcinomas, pS9GSK3╬▓ expression is higher in AC than in other subtypes such as SCC, large cell carcinoma, and small cell carcinoma [11], suggesting the possible association of pS9GSK3╬▓ expression with the histological phenotype of AC. However, in ACs of organs other than lung and uterine cervix such as in gastric and colorectal ACs, the data on expression levels of pS9GSK3╬▓ are conflicting, suggesting its organ specificity [8,10].
Previously, we have shown that there is significant overexpression of GSK3╬▓ in SCC than in AC, suggesting the association of GSK3╬▓ with the squamous phenotype in cervical cancer [13]. In this study, we have not only verified the previous results but have also shown a possible association between membranous expression of pS9GSK3╬▓ and cervical AC. The result of this study suggests that different expression pattern of GSK3╬▓ and pS9GSK3╬▓ is associated with the histologic phenotype of cervical cancer. This also implies that extreme caution needs to be taken while using GSK3 inhibitor in cervical cancer patients, especially in AC patients, because it may lead to unintentional cancer cell growth and proliferation [30].
In conclusion, the phosphorylation status of GSK3╬▓ changes during cervical cancer development and the differential expression patterns of GSK3╬▓ and pS9GSK3╬▓ may be associated with the histologic phenotype of cervical cancer, possibly, GSK3╬▓ with SCC and pS9GSK3╬▓ with AC.






")











